| |

La falconeria moderna è molto cambiata nelle
tecniche e nelle strumentazioni usate rispetto alla
falconeria di un tempo. Oggi si usano solo rapaci
riprodotti in cattività, si usano aquiloni e palloni
aerostatici ad elio per addestrarli, se si perdono
si ricorre ad apparecchiature da radiotracking e le
specie stesse di rapaci utilizzate sono diverse. Ai
classici Pellegrini, Sacri, Lanari, Astori e
Sparvieri oggi si aggiungono specie quali la Poiana
di Harris (Parabuteo unicinctus) o la Poiana
codarossa (Buteo jamaicensis) che provengono
dal continente americano ma si tratta ancora di
specie “naturali” per così dire. Il fatto è che oggi
si usano in falconeria anche specie, diciamo
“artificiali”, cioè gli ibridi che, in Italia vanno
e in qualche altro Paese stanno andando molto di
moda.
I primi ibridi che si produssero nei lontani anni
’70 in USA servivano più per scopi di ricerca
biologica che altro (Baptiste, S. and D. Jamieson. 1978,
Cade, T.J. and J.D. Weaver. 1976, Boyd, L. and N.
Boyd. 1975),
ma poi anche i falconieri e gli allevatori privati
si appropriarono delle necessarie tecniche di
inseminazione artificiale e di ibridologia ed
iniziarono così una produzione in proprio di tali
esemplari per l’uso pratico in falconeria.
GIA DEGLI IBRIDI
Gli ibridi nascono da genitori di Specie diversa. In
nessun caso, sia dall'incrocio fra ibridi, che da
qualunque loro reincrocio, si ottiene una loro
genealogia, cioè una nuova Specie ma, al massimo, si
può ottenere una “Razza”. Per capire i
fondamenti dell’ibridologia dei rapaci bisogna
considerare alcuni concetti fondamentali, che
sarebbe meglio illustrare con degli esempi basati su
specie più note. Iniziamo con il Canarino.
Fin dai primi anni della sua introduzione in Europa,
il Canarino è stato incrociato con Specie affini
europee ed extraeuropee, con risultati in molti casi
positivi. Però nessun prodotto di questi incroci,
cioè nessuno di questi ibridi si è perpetuato, a
conferma che la Natura tende a mantenere le Specie
in purezza e concede solo il perpetuarsi degli
incroci fra individui della stessa Specie (meticci),
anche se appartenenti a Razze esteriormente assai
differenti fra loro; si pensi che nell'ambito della
Specie Canina, qualunque incrocio è fecondo, anche
quello fra un gigantesco Alano ed un minuscolo
Yorkshire!
LASSIFICAZIONE DEGLI IBRIDI
Dunque un ibrido è l’incrocio tra due specie
diverse, ma bisogna chiarire che si possono avere
tre tipi di ibridi: a) incroci, cioè accoppiamenti
tra esemplari della stessa sottospecie ma di ceppo
diverso o comunque con caratteristiche diverse
(sotto certi punti di vista in questo caso non si
potrebbe parlare di veri e propri ibridi, ma appunto
di incroci tra individui con caratteristiche diverse
o appartenenti a ceppi diversi; nei rapaci un
esempio potrebbe essere l’incrocio tra diverse fasi
di colore, visto che ancora la selezione artificiale
non ha portato alla creazione di veri e propri
ceppi); b) ibridi intraspecifici (cioè tra due
sottospecie o razze della stessa specie per es.
Falco peregrinus brookei x Falco peregrinus
peregrinus); ibridi interspecifici o
intragenerici (cioè tra due specie appartenenti allo
stesso genere per es. Falco peregrinus x Falco
rusticolus); c) ibridi intergenerici (cioè fra
generi diversi per es. tra Poiana codarossa,
Buteo jamaicensis e Poiana di Harris,
Parabuteo unicinctus oppure tra Sparviere di
Cooper Accipiter cooperii e Poiana di
Harris).
Si possono ottenere anche degli ibridi di seconda
generazione (F2) cioè quelli ottenuti dall’ incrocio
di un ibrido (per es. Girfalco x Lanario) con una
specie pura (per es. Sacro) per ottenere così un
ibrido finale Girfalco/Lanario x Sacro nel caso
degli ibridi eterologhi (cioè reibridazione con una
specie esterna), mentre se otteniamo in F2 ibridi
del tipo Girfalco/Sacro x Sacro si parlerà di
omologhi, cioè derivati dall’incrocio di un genitore
ibrido (nel nostro esempio il Girfalco x Sacro) con
un genitore appartenente ad una delle due specie
parentali (pure) da cui era stato ottenuto l’ibrido
F1 (nel nostro esempio il Sacro); nel caso
dell’ottenimento di ibridi F2 omologhi l’incrocio
usato è detto back-crossing (cioè retroincrocio,
appunto perché abbiamo riaccoppiato l’ibrido con una
delle due specie parentali da cui esso proviene e si
indica con R, quindi la prima generazione di
reincrocio si indica con R1, la seconda con R2 e
così via); i suddetti ibridi omologhi ed eterologhi
sono più comunemente chiamati triibridi. Ma in F2 si
possono avere anche i così detti ibridi
bi-eterologhi (in realtà la nomenclatura dovrebbe
essere più complessa…) cioè derivati dall’incrocio
tra due genitori entrambi ibridi (per esempio
Girfalco/Sacro x Girfalco/Lanario); tali ibridi
vengono più comunemente chiamati quadriibridi.
La nomenclatura appena citata, di tri-ibridi e
quadri-ibridi in realtà non è del tutto corretta.
Come si può notare da quanto detto sopra gli ibridi
possono essere classificati anche quantitativamente
cioè semplicemente in base solo alle specie che ci
sono dentro: si parlerà allora di bi-ibridi quando
si ottiene un ibrido da due specie pure diverse
oppure quando viene reincrociato un ibrido F1 con
una delle due specie pure che lo compongono (GyrxLan
è un bi-ibrido, ma anche Gyr/Lan x Lan lo è perché
anch’esso è composto da due specie anche se
combinate in percentuali diverse con un reincrocio);
si parla di tri-ibridi quando le specie coinvolte
sono tre, di solito nel caso degli ibridi F2
eterologhi come ad esempio il Gyr/Lan x Sacro; ed
infine si parla di quadri-ibridi quando le specie
coinvolte sono quattro; i quadri-ibridi sono
rarissimi perché possono essere ottenuti solo da
incroci di due genitori a loro volta ibridi, ma per
quanto diremo dopo, le femmine ibride F1 sono di
solito sterili o poco fertili per cui le difficoltà
di produzione di un Gyr/Sacro x Lan/Lugger per
esempio sono notevolissime.
Prima di continuare è bene chiarire il sistema di
nomenclatura degli ibridi: si mette prima la specie
del genitore di sesso maschile (il padre) e poi la
specie del genitore femminile (la madre); allora nel
caso di un ibrido Girfalco x Lanario capiremo subito
che questo individuo è figlio di padre Girfalco e di
madre Lanario. Nel caso di bi-ibridi (cioè di ibridi
di prima generazione derivati dal semplice incrocio
tra maschio e femmina di specie diverse) si usa un
segno “x” per collegarli (per esempio Girfalco x
Lanario o, abbreviato GirxLan); nel caso invece di
ibridi di seconda generazione, siano essi omologhi
od eterologhi, per descrivere l’esatta composizione
della F2 ottenuta si userà il segno “/” per legare
le due specie da cui proviene uno dei genitori o
entrambi (nel caso degli ibridi bi-eterologhi) ed il
segno “x” per rappresentare l’ulteriore incrocio tra
essi (per esempio: un ibrido eterologo può essere
Girfalco/Pellegrino x Sacro mentre un bi-eterologo
può essere Girfalco/Sacro x Girfalco/Lanario).
Un ulteriore fattore di classificazione degli ibridi
è la direzione dell’ibrido. Vediamo prima un
esempio. Un esempio di ibrido fra i più noti è il
Mulo che nasce da Cavallo x Asina, o viceversa se si
incrocia Asino x Cavallo si ottiene il Bardotto che
è abbastanza diverso dal mulo per costituzione
fisica e per comportamento. Applicando ai rapaci
possiamo dire che un GirxPell sarà abbastanza
diverso da un PellxGir, anche se questo non è
facilmente dimostrabile nella pratica, ma ci sono
delle spiegazioni: un allevatore che produce ibridi
di rapace userà quasi esclusivamente l’inseminazione
artificiale (gametizzazione artificiale) e su questo
non c’è alcun dubbio, ora l’allevatore, dovendo
produrre degli ibridi tra la specie Girfalco e la
specie Pellegrino (molto rinomati in falconeria, e
costosi), preferisce usare un maschio di Girfalco da
cui prelevare il seme ed una femmina di Pellegrino
da inseminare col seme del Girfalco, semplicemente
per una questione economica, infatti acquistare un
maschietto di Girfalco costerà sicuramente meno che
acquistare una femmina, mentre tra femmina e maschio
di Pellegrino la differenza di prezzo è irrisoria;
ecco perchè gli ibridi tra la specie Pellegrino e la
specie Girfalco che troviamo in giro sono quasi
tutti di direzione GirxPell e non viceversa
PellxGir. Questo è uno dei motivi, ma ce ne sono
anche altri, troppo complessi, che non mi dilungo a
spiegare, anche perchè io stesso non ne ho perfetta
conoscenza.
Quello che segue è, solo a livello di esempio, un
breve elenco solo di alcuni tipi di ibridi che
possono essere ottenuti da sole 3 specie, in
funzione dei concetti sopra esposti. Si consideri
che questi ibridi sono tutti diversi tra loro
(etologicamente, biologicamente e morfologicamente)
al di là della variabilità individuale:
Specie
considerate: Falco sacro, Falco lanario e Girfalco
1) GyrxLan
2) GyrxSacro
3) LanxGir
4) LanxSacro
5) SacroxLan
6) SacroxGir
7) Gyr/Lan x Sacro
8) Gyr/Lan x Lanario
9) Gyr/Lan x Girfalco
10)
Gyr/Sacro x Sacro
11)
Gyr/Sacro x Lanario
12)
Gyr/Sacro x Sacro
13)
Lan/Gyr x Girfalco
14)
Lan/Gyr x Lanario

Variabilità di colore degli ibridi tra Girfalco e
Sacro

Ibrido Girfalco nero x Sacro
LE PROPORZIONI DEGLI IBRIDI E LA QUESTIONE DEL
REINCROCIO
Come è facile capire, in un ibrido F1, poniamo un
GirxLan, troveremo teoricamente (si veda dopo per la
spiegazione del “teoricamente”) il 50% di geni del
Lanario ed il 50% di geni del Pellegrino. Quindi
tutti gli ibridi F1 avranno un rapporto genetico
teorico del 50:50 %. In un ibrido F2 se questo è
omologo, cioè per es. se il GyrxLan viene
reincrociato con un Lan o con un Gyr, la progenie
ottenuta avrà rispettivamente il 75% di geni dal
lanario ed il 25% dal Girfalco nel primo caso (i
Gyr/Lan x Lan sono i così detti ¾ Lanario) o il 75%
di geni dal Girfalco ed il 25% dal Lanario nel
secondo caso (i Gyr/Lan x Gyr sono i così detti ¾
Gyr cioè tre quarti Girfalco; ma teniamo conto che
un ¾ Gyr può essere anche il Gyr/Sacro x Gyr). Se
invece l’ibrido è eterologo le proporzioni saranno
diverse; per esempio se incrociamo il GyrxLan con un
Sacro i tri-ibridi (eterologhi) che otteniamo nella
F2 avranno il 25% di geni dal Lanario, un altro 25%
di geni dal Girfalco ed il 50% di geni dal Sacro.
Ma le proporzioni sopra illustrate sono solo
teoriche, perché sebbene un ibrido abbia una
percentuale di geni da un genitore e il resto
dall’altro non è detto che manifesti questi geni nel
suo fenotipo allo stesso modo. Un ibrido GyrxLan che
avrà quindi il 50% teorico di geni dal Girfalco ed
il rimanente 50% teorico dal Lanario, può apparire
nella realtà (nel suo fenotipo) molto più simile al
Girfalco che non al Lanario. Ciò avviene per vari
motivi: intanto perché i geni si rimescolano
nell’individuo (attraverso il meccanismo del
crossing-over) e poi perché i geni interagiscono
fortemente tra loro, per cui un gene che nella
specie pura si manifesta, nell’ibrido potrebbe
essere inibito (epìstasi) da uno o più geni
dell’altra specie e quindi non si manifesta o si
manifesta in maniera diversa. Volendo approfondire
in termini più tecnici questo ultimo concetto
bisogna introdurre il concetto di coadattamento
genetico. I geni interagiscono non solo con
l’ambiente ma anche con gli altri geni presenti
nell’organismo. Per ogni locus genico la selezione
naturale favorisce gli alleli che interagiscono bene
con gli alleli negli altri loci. Il termine
coadattamento genetico si riferisce all’interazione
di tipo adattativo tra i geni che costituiscono un
pool genico. E la non vitalità o la sterilità degli
ibridi interspecifici rappresenta infatti una
notevole prova del coadattamento genetico (per es. i
genotipi del cavallo e dell’asino non sono
reciprocamente adattati e la conseguenza è infatti
che i loro ibridi non sono fertili). Ma il
coadattamento genetico influenza anche le
proporzioni fenotipiche che si manifestano negli
ibridi, perché se due genotipi non sono coadattati
(come lo sono quelli delle due diverse specie pure
da cui si ottiene l’ibrido), i loro geni
interagiranno in maniera anomala e di solito
imprevedibile per cui quello che otterremo
fenotipicamente nell’ibrido sarà il risultato di
interazioni “forzate” tra geni anche non compatibili
e che dunque potranno anche non esprimersi, o farlo
solo parzialmente o in modo anomalo.

Tri-ibrido omologo 3/4 Girfalco bianco x Sacro

Tri-ibrido eterologo GirfalcoxLanario/Sacro
In tutti questi rapporti genetici però esiste una
piccola questione che potrebbe portare a confusione
per cui ve la mostro nella speranza di riuscire a
spiegare adeguatamente il concetto, che in realtà è
abbastanza complesso. Infatti, a proposito degli
ibridi tra Cavallo e Asino, in alcuni testi si legge
che il Mulo (che è un ibrido di Cavallo x Asina) è
sempre fertile se di sesso femminile e se viene
reincrociato (back-crossing) con un Cavallo darà
Cavalli e Muli, mentre se viene reincrociato con un
Asino darà come progenie Asini e Muli. Si deduce
questo dal seguente schema (dove indicando con CC il
cavallo e con AA l’asino si ha che CA è il Mulo
mentre AC è il Bardotto):
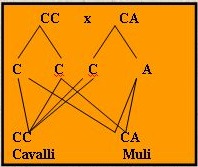
Come si vede dal grafico sopra, in questo tipo di
incroci le proporzioni genetiche non si sommano ma
si mantengono distinte e si ritorna nuovamente alle
specie parentali con la progenie ottenuta dal
reincrocio (infatti praticamente si riottengono il
50% di cavalli puri ed il 50% di Muli). Allora,
seguendo questo ragionamento dal reincrocio di
GyrxLan con una femmina Gyr si dovrebbero ottenere
come figli il 50% di Girfalchi puri ed il 50% di
ibridi GyrxLan (per esempio 2 Girfalchi e 2
GyrxLan) piuttosto che ottenere il così detto ¾ Gyr!
Come tutti sappiamo la realtà sembra non essere così
e infatti non è così. A questo proposito si fa una
distinzione tra Ibridi mendeliani (che sono quelli
dell’esempio con il Mulo) perché seguono gli stessi
principi usati da Gregor Mendel per la dimostrazione
delle sue leggi sull’ereditarietà dei caratteri, nei
quali le proporzioni genetiche si mantengono
separate, ed ibridi non mendeliani o quantitativi
(che sono quelli degli esempi sulle frazioni
riportati di sopra), nei quali le proporzioni
genetiche sono additive cioè si sommano di volta in
volta andando contro le teorie di Mendel, tanto è
vero che si sa che ci sono alcuni caratteri (i
caratteri quantitativi) che non rispettano le leggi
mendeliane proprio perché sono additivi. Questo
almeno in teoria, perché verificando nella pratica
si vede che è impossibile che i geni di due specie
diverse che si trovano in un ibrido possono rimanere
completamente separati senza interagire tra loro
(crossing-over) per cui è impossibile che si possa
poi ritornare con il reincrocio alla specie pura,
resterà sempre un residuo di geni, che calerà tanto
di più quanto più volte viene reincrociato l’ibrido
iniziale con la specie pura, riducendo così la
frazione di geni dell’altra specie (per es. R1
50:50, R2 25:75, R3 12,5:87,5 ecc.) fino ad arrivare
ad una riduzione infinitesimale della percentuale
del genoma della specie estranea. Possiamo quindi
concludere che praticamente gli ibridi mendeliani
non esistono! Questo perché Mendel lavorava con dei
singoli alleli (l’allele è una delle due copie che
ogni gene ha negli organismi superiori che sono
appunto diploidi) mentre negli ibridi si lavora con
dei genomi interi. Mentre il singolo gene agisce in
maniera semplice invece un genoma intero (cioè l’
insieme dei geni di una specie) opera in maniera
estremamente complessa (coadattamento gentico per
es.), ecco perché il grafico precedentemente visto
non è valido; in esso quelli che si incrociano sono
degli interi genomi (quelli del cavallo e del mulo,
che a sua volta ha un genoma ibrido tra cavallo e
asino) che dunque ben difficilmente potranno tornare
al loro stato puro, mentre per un singolo allele è
più facile spostarsi, nelle generazioni dalla forma
eterozigote a quella omozigote. Detto questo
possiamo affermare quindi che il concetto secondo
cui il reincrocio (R1) tra mulo e cavallo dia 50% di
muli e 50% di cavalli non corrisponde a verità.
Quelli ottenuti saranno invece dei ¾ Cavallo (e ¼
Asino), in cui magari alcuni individui somigliano di
più a Cavalli puri (ma non lo sono!) e altri
somigliano di più ai muli. FERTILITA’ DEGLI IBRIDI
Si osserva a questo proposito un altro fenomeno
molto interessante: il Mulo maschio è sempre sterile
(arresto della spermatogenesi alla fase di
spermatocita primario), mentre la Mula è spesso
fertile, sia con l'Asino che con il Cavallo; ciò
esclude la possibilità che si formi una genealogia
di soli Muli, cioè una nuova Specie. Tale differenza
rispetto al Mulo e ad altri ibridi di Mammiferi (in
questi sono i maschi sempre sterili) ha fatto capire
facilmente che il sesso sterile è quello
eterogametico (regola di Haldane). Nei
Mammiferi la coppia di cromosomi sessuali XY (sesso
eterogametico) spetta ai maschi, negli Uccelli alla
femmina. Applicando ai rapaci infatti è risaputo che
gli ibridi fertili sono i maschi e più difficilmente
le femmine (ecco perchè è difficile produrre gli
ibridi bi-eterologhi) e allora nella produzione di
ibridi F2 si preferisce scegliere la direzione
Gir/Lan x Sacro (cioè il padre ibrido e la madre
pura) e non Sacro x Gir/Lan cioè il padre puro e la
madre ibrido, appunto perchè le femmine ibride hanno
sempre un elevato grado di sterilità, anche se non
assoluta.
Altro importantissimo fattore che condiziona la
fertilità degli ibridi è la loro sistematica: in
base ai rapporti filogenetici tra essi intercorrenti
si può già capire se è possibile o meno ottenere un
determinato ibrido. Tali concetti sono illustrati
nel paragrafo seguente.SISTEMATICA DEGLI IBRIDI
Oggi la maggior parte degli ibridi prodotti
artificialmente appartengono al genere Falco. Alcune
di queste specie hanno corredi cromosomici
completamente differenti ed i loro ibridi sono
probabilmente infertili. Il genere Falco oggi è
stato suddiviso in generi e sottogeneri come per
esempio il gruppo dei Pellegrini (Rhincodon)
e quello dei grossi falconi o falchi del deserto
(Hierofalco); gli ibridi fra questi sottogeneri
sono di solito molto “incrociati” e sono meno
fertili degli ibridi ottenuti all'interno di ciascun
sotto genere, proprio perchè il grado di diversità è
notevole. Così un ibrido tra Falco sacro e Falco
pellegrino è più “incrociato” rispetto ad un ibrido
tra Falco sacro e Girfalco o tra "Shaheen" e
Pellegrino. Le ricerche sugli ibridi stanno ancora
continuando soprattutto per chiarire la relazione
tra il Girfalco ed il Falco sacro, infatti, è facile
che queste due specie siano una singola specie. Essi
sono completamente fertili anche dopo varie
generazioni per cui si inizia a dubitare del fatto
si possa parlare di ibridi. Da quanto detto finora
si capisce allora perché gli ibridi con il
Pellegrino si fermino spesso alla prima generazione
(F1). Un ibrido PellxLan infatti è spesso infertile
e non potrà dare progenie se reincrociato con un
Lanario, forse potrebbe in alcuni rari casi dare una
prole F2 se reincrociato di nuovo con un Pellegrino.
Se ci riflettete bene la maggior parte dei di
tri-ibridi F2 conosciuti sono sempre tra Lanario,
Sacro e Girfalco (Gir/LanxSacro oppure Gyr/SacroxLan
o anche Gyr/SacroxSacro ecc.), che, appunto
appartengono allo stesso sottogenere (Hierofalco,
Falchi del deserto). Che certi incroci fra Cane e
Lupo, o fra Muflone e Pecora, o fra Germano reale e
Anatra domestica, diano prole regolarmente feconda
in entrambi i sessi, non costituisce eccezione,
perché i casi di questo genere non sono altro che
incroci fra il progenitore selvatico e il suo
derivato domestico: perciò si tratta di specie ad
elevata affinità genetica. In effetti è tutta
questione di affinità, ma non di affinità delle
forme esteriori, bensì dei corredi cromosomici dei
due "partners", e si capisce subito perché
l'interpretazione di questi fenomeni sia molto
complessa e difficile: perché studiare e capire
questi cromosomi degli ibridi, già dal punto di
vista della manualità sperimentale è estremamente
arduo. La pratica dell'ibridismo riveste allora un
ruolo fondamentale nella Sistematica, perché è
ancora uno dei criteri più validi per stabilire se
due individui appartenenti a popolazioni differenti
debbano essere ascritti a Specie differenti, o alla
stessa Specie (ricordiamo che per la classificazione
degli Uccelli sono state utilizzate tecniche
molecolari di ibridazione diretta dei DNA di due
specie, quelle usate da Sibley per es. per studiare
la sistematica degli Uccelli). Di regola, se due
individui, anche apparentemente diversissimi fra
loro, danno prole feconda, si stabilisce che
appartengono alla stessa Specie (in tal caso la
differenza è di Razza o Sottospecie); se la prole
nasce, ma è infeconda, si tratta di due Specie
differenti appartenenti allo stesso Genere
(differenza di Specie). Se non si conoscono casi di
nascita di prole, si stabilisce che il Genere è
differente, anche se altri caratteri autorizzano a
far appartenere i due individui alla stessa Famiglia
(ma per questo ultimo caso ci sono molte eccezioni).
Per quando riguarda i rapaci però questa regola
apparentemente fa acqua da tutte le parti. Da quanto
si è visto dai risultati delle ibridazioni, infatti,
gli ibridi interspecifici ottenuti sono di solito
fertili, e si possono ottenere anche gli F2, ma
questo può essere spiegato con la classificazione
precedente del genere Falco in Sottogeneri ad alta
affinità interna. Inoltre nei rapaci si conoscono
anche ibridi intergenerici ad esempio quello tra
Poiana di Harris, Parabuteo unicinctus e
Astore Accipiter gentilis, o tra Poiana di
Harris, Parabuteo unicinctus e Poiana
codarossa , Buteo jamaicensis, in questo
caso la spiegazione è filogenetica, infatti il
genere Parabuteo comprende al suo interno una sola
specie (P. unicinctus) che per una serie di
caratteristiche possedute è stata classificata a
parte (non è Buteo né Accipiter) ma il fatto che dia
ibridi si a con il genere Buteo che con il genere
Accipiter deve far riflettere sulla sua corretta
posizione sistematica.
|
SOTTOGENERE |
SPECIE |
|
Rhyncodon |
peregrinus
pelegrinoides
kreyenborgi |
|
Hierofalco |
rusticolus
cherrug
biramicus
jugger
mexicanus,
subniger (?)
hypoleucos (?) |
|
Nesierax |
deiroleucus
rufigularis
femoralis
novaesaelandiae |
|
Aesalon |
columbarius
chiquera (?) |
|
Hypotriorchis |
fasciinuncha (?)
subbuteo
cuvieri
severus
longipennis
eleonorae
concolor
vespertinus
amurensis,
|
|
Tinnunculus |
tinnunculus
moluccensis
cenchroides
sparverius
newtoni
punctatus
araea
naumanni
alopex
rupiculoides |
|
Gheppi aberranti |
ardosiaceus,
dickinsoni,
zoniventris, |
|
Hieracidae |
berigora |
Filogenesi del Genere Falco (mod. da N. Fox, tesi di
laurea, Canterbury University, 1977).
Le
specie sono naturalmente adattate a mantenersi
uniformi, cioè a non produrre ibridi. Per capire
meglio dobbiamo rifarci al concetto di specie: sono
state date tantissime definizioni della “specie”,
alcune più valide di altre (Mayr, Lisley, Usinger,
1953, Simpson, 1968, Wiley, 1978, Paterson, 1985).
Definire la specie comunque rimane difficile:
cercando di sintetizzare tutte le varie definizioni
date si potrebbe dire che la specie può essere vista
come un complesso di popolazioni naturali la cui
unità deriva dalla sua origine monofiletica; tale
unità si mantiene nei limiti spazio-temporali nel
cui ambito le sottounità discrete che in ogni
momento lo formano ne mantengono la coesione
riproduttivo-genetica interna e l’indipendenza del
pool genico; in conseguenza di ciò la specie
interagisce in modo unitario con l’ambiente. In
altre parole la specie è una popolazione costituita
da individui geneticamente simili (unità della
popolazione) perché si sono originati da uno stesso
progenitore comune (origine monofiletica); questa
popolazione riesce a mantenersi uniforme e unita nel
tempo e nello spazio perché tutti i vari individui
che la compongono (le sottounità discrete) sono
geneticamente isolate cioè non in grado di
accoppiarsi con altri individui di altre “specie” e
dando ibridi fertili.
In
base alla definizione data possiamo dire che un
ibrido è la mescolanza di geni “alieni” con il pool
genetico di una popolazione mendeliana (Sybley,1957,
Rising, 1983). L’ibrido è dunque la progenie di un
incrocio tra due individui o popolazioni
geneticamente diverse (con un certo grado di
diversità).
Dunque
è questa coesione riproduttivo-genetica degli
individui di una specie che ostacola la formazione
di ibridi. Però se teniamo conto che due specie
possono essere molto simili perché a loro volta
hanno avuto una origine evolutiva da un progenitore
comune, allora possiamo avere qualche speranza di
riuscire a produrre degli ibridi. Ecco perché, per
esempio, grazie alla produzione di ibridi, ma anche
grazie ad altre osservazioni, si è compreso che il
Lanario, il Girfalco, il Sacro, il Lugger, sono
strettamente imparentati tra loro, anche se non a
tal punto da essere considerati tutti un’unica
specie.
In
ogni momento sul Pianeta nascono nuove specie e se
ne estinguono altre, in maniera naturale e a
prescindere dall’influenza dell’uomo. I zoologi
sistematici hanno sviluppato varie teorie per
spiegare la speciazione (cioè la nascita naturale di
una nuova specie) ed una delle teorie oggi
abbastanza confermata è quella della speciazione per
ibridazione. Si conoscono infatti attualmente
diverse specie ormai isolate (e con tutte le
caratteristiche tipiche di una “specie” secondo le
definizioni date prima) che sono nate da ibridazione
continua e ripetuta tra due specie diverse (un
esempio è quello di Rana esculenta = Rana
lessonae x Rana ridibunda) . Anche nei
rapaci si ha un caso del genere: infatti si è
ipotizzata la teoria della speciazione per
ibridazione allo scopo di spiegare l’origine e la
filogenesi del Falco degli Altai (Falco altaicus),
un rapace la cui popolazione vive esclusivamente
nelle montagne dell’Asia centrale e che ha delle
caratteristiche particolari che lo distinguono dalle
altre popolazioni di Falco Sacro e allo stesso tempo
simile ad alcune popolazioni di Girfalco ma
sostanzialmente diverso anche da questo. Su questa
specie sussiste un dibattito ancora aperto (cfr. per
es. Ellis, 1995): esiste il Falco altaicus? O
sarebbe meglio considerarlo una ssp. del Falco
sacro, e quindi Falco cherrug altaicus?
Alcuni, hanno allora proposto la teoria della
speciazione per ibridazione, affermando che il “Falco
altaicus” può essere considerato come una specie
a se stante e derivata da ibridazione tra Falco
sacro e Girfalco (che appunto, da quanto si è visto
in cattività, sono in grado di produrre ibridi
fertili anche per diverse generazioni).
OTTENIMENTO DEGLI IBRIDI
Come
abbiamo visto, la prima difficoltà che si incontra
nella produzione di ibridi è la sterilità, che è
dovuta sia alla loro posizione sistematica sia al
loro sesso (Legge di Haldane). Quando, inoltre, nel
campo dell'ibridismo, si parla delle difficoltà
riproduttive, occorre distinguere fra le difficoltà
che si incontrano nell'ottenere gli ibridi e le
difficoltà che si incontrano nell'ottenere prole dal
reincrocio dell'ibrido con la specie di uno dei due
genitori nell'intento di ottenere gli R1, R2, ecc.
Questo perché nel primo caso (ottenere gli ibridi da
due specie pure) ci sono delle difficoltà
intrinseche più legate alla tecnica di produzione
che alla loro sterilità (posto che stiamo lavorando
con specie “compatibili” cioè abbastanza affini,
perché parlare di ibridi tra Aquila reale e Falco
pellegrino vi renderete conto che è
fantascientifico, oltre che inutile). Nel secondo
caso (ottenere ibridi F2 da due ibridi o da un
ibrido ed una specie pura) alle difficoltà tecniche
di produzione si aggiunge anche il problema della
sterilità degli ibridi.
Alla
nascita di ibridi si oppongono vari fenomeni
biologici dovuti all’adattamento, poiché gli ibridi
nelle popolazioni selvatiche, solitamente hanno una
fitness più bassa. Oltre alle così dette barriere
riproduttive fisiologiche (incompatibilità del
corredo cromosomico, degli spermatozoi con le uova
ecc.) esistono anche degli adattamenti
comportamentali che inibiscono l’accoppiamento di
due individui di specie diverse (barriere
etologiche): l’imprinting sui genitori (parent
imprinting) è il principale.
Per la
produzione di ibridi artificiali allora la tecnica
principale è l’inseminazione artificiale
(gametizzazione artificiale) che permette di
superare gli ostacoli comportamentali ma non le
barriere fisiologiche prima citate. La prima cosa da
fare è allora quella di scegliere individui di
specie compatibili, cioè abbastanza simili
filogeneticamente da non opporre resistenza
all’incrocio (gli spermatozoi di Girfalco allora,
incontreranno poca resistenza a penetrare in una
cellula uovo di Lanario per fecondarla, mentre ne
incontreranno di più per fecondare una cellula uovo
di Pellegrino e saranno probabilmente incapaci di
fecondare una cellula uovo di Astore). Come avevo
accennato precedentemente oggi gli ibridi sono
prodotti soprattutto a scopo di falconeria, la
scienza non ha più bisogno di ibridare gli individui
per studiarne i rapporti filogenetici, perché può
ricorrere alle tecniche molecolari di ibridazione
diretta del DNA di specie diverse o addirittura di
confronto, nucleotide per nucleotide, attraverso il
sequenziamento di piccoli pezzi di DNA di due
specie. Gli allevatori privati che producono ibridi
per la falconeria, lo fanno a scopo economico
soprattutto e dunque preferiranno ad es. usare un
maschio di Girfalco piuttosto che una femmina (più
costosa) per produrre ibridi GirxPell. Allo stesso
modo è sempre l’aspetto economico che ci può dare
una spiegazione a tutti i vari tri- e quadri-ibridi
(omologhi o eterologhi) prodotti finora.
Alcuni
allevatori però non vogliono cimentarsi con
l’inseminazione artificiale e sfruttano una tecnica
più semplice ma meno produttiva: l’imprinting
parentale incrociato (cross-imprintig). Come abbiamo
detto prima l’imprinting sui genitori influenza i
comportamenti inter- ed intra-specifici
dell’individuo e le sue scelte sessuali (mate
selection o sexual selection). Se un dato animale
viene allevato dai suoi genitori naturali, o da
genitori comunque della sua stessa specie, esso
imparerà nelle sue prime settimane di vita a
riconoscere i suoi con specifici e da adulto, quando
sarà sessualmente maturo, cercherà un partner per la
riproduzione della stessa specie dei suoi genitori e
cioè della sua stessa specie. Se, viceversa viene
allevato da genitori di specie diversa, per es.
quando un Lanario viene allevato da genitori
Girfalco, esso da adulto riconoscerà nei Girfalchi i
suoi conspecifici e tenterà di accoppiarsi con un
partner Girfalco. In quest’ultimo caso se anche il
partner Girfalco sarà stato allevato da genitori
Lanario, entrambi si riconosceranno a vicenda come
della stessa specie e, compatibilmente con i loro
patterns etologici istintivi (parate nuziali, che
nei Falchi sono molto simili) si corteggeranno e si
accoppieranno deponendo uova fertili ed allevando
una nidiata di ibridi GirxLan. Il cross imprintig (
o imprinting incrociato ) è questo e garantisce in
teoria al 100% la possibilità di ottenere ibridi,
ma, si sa, con i rapaci è già complesso ottenere una
riproduzione naturale, per cui per una ibridazione
naturale le probabilità calano. Comunque è un metodo
efficace anche se l’inseminazione artificiale
garantisce risultati migliori.
Nel
caso dell’ottenimento di ibridi F2 il
cross-imprinting verrà applicato allo stesso modo,
cioè se vogliamo ottenere ibridi F2 da padre GyrxLan
e madre Lanario (per ottenere dunque degli ibridi
Gyr/Lan x Lanario) dovremo fare allevare i nidiacei
GyrxLan da genitori Lanari, in modo tale che essi da
adulti riconoscano come conspecifici i Lanari e li
possano scegliere come loro partner per la
riproduzione. Se ovviamente gli ibridi sono stati
allevati a mano (come spesso avviene negli
allevamenti privati, sempre per un motivo economico)
essi saranno imprintati con l’uomo e difficilmente
accetteranno di accoppiarsi spontaneamente con altre
specie di falchi (anche se i casi di re-imprinting
cioè di ridirezione dell’imprinting sessuale verso
un’altra specie sono abbastanza frequenti); per essi
sarà meglio usare la gametizzazione artificiale
cooperativa.
Il
fenomeno del cross imprinting spiega anche perché un
eventuale ibrido sfuggito alla cattività, ben
difficilmente tenterà di accoppiarsi con un rapace
selvatico a meno che l’ibrido (poniamo un GirxLan)
non sia stato esso stesso allevato da genitori
diversi, poniamo Lanari; in tal caso l’ibrido
sfuggito alla cattività potrebbe tentare di
accoppiarsi con un Lanario selvatico, ma poiché
sicuramente il Lanario selvatico è stato allevato
dai suoi genitori Lanari, esso nella maggioranza dei
casi non accetterà di accoppiarsi con l’ibrido, che
non riconoscerà come un suo conspecifico. ILITA’ DEGLI IBRIDI E LORO APPLICAZIONI PER
LA FALCONERIA MODERNA
Fortunatamente oggi nella maggioranza dei Paesi è
vietata la cattura dei rapaci selvatici a scopo di
falconeria. Ma la falconeria non è morta perché
alcuni falconieri si sono impegnati, per primi,
nella riproduzione in cattività dei rapaci,
principalmente di quelle specie più comunemente
usate. Però i ceppi in cattività non erano molto
grandi per cui era facile incrociare anche
sottospecie diverse di falchi pur di ottenere una
progenie. Questi rapaci che si riescono a produrre
in cattività, incrociati o puri, comunque riescono a
soddisfare le esigenze dei falconieri. Ma già a
partire dagli anni ’70 molti falconieri e
ricercatori si cimentarono nell’applicazione delle
tecniche di inseminazione artificiale dei rapaci.
Tale tecnica aprì molte prospettive per la ricerca e
la conservazione (perché permette di riprodurre
anche individui restii a riprodursi naturalmente per
es. rapaci irrecuperabili e non più rilasciabili in
natura) ma anche per i falconieri che la
utilizzarono e la utilizzano tuttora per produrre
degli ibridi, un nuovo modo di guardare all’arte
della falconeria.
Ma
perché si vogliono produrre gli ibridi e quali sono
i loro vantaggi? Gli ibridi esistono anche in molte
altre specie animali molto note quali cani, cavalli,
gatti, ecc…Negli animali e nelle piante domestiche
molte razze sono state selezionate proprio con i
metodi dell’ibridazione ed in essi la così detta
eterosi dell’ibrido ha portato ad un incremento
nelle dimensioni, nella rusticità e quindi anche
nella resistenza alle malattie. Tutti questi animali
assieme anche agli uccelli da gabbia sono stati
ibridati allo scopo di far risaltare determinati
caratteri quali la bellezza, la robustezza, il canto
ecc..; comunque alla fine qualsiasi sia la
motivazione, l’animale finale ibrido, poiché sarà
composto da vari elementi, produrrà molti dei
fattori che si desiderava ottenere. Nell’
ibridazione dei falconi i caratteri che si desidera
ottenere sono il colore, la dimensione, la potenza,
l’altezza di volo e l’ attitudine ad inseguire e
catturare una preda. E molti ibridi, infatti,
presentano queste caratteristiche. Quindi il motivo
principale dell’ibridazione dei rapaci a scopo di
falconeria è la produzione di esemplari che
possiedono delle nuove caratteristiche in grado di
soddisfare tutte le esigenze della falconeria
moderna.
Bisogna però dire che non tutte le ciambelle escono
col buco, così nella produzione di ibridi se da un
lato si ottengono esemplari che veramente assommano
positivamente le caratteristiche di due specie
diverse, dall’altro si possono ottenere anche
esemplari che assommano le caratteristiche negative
delle due specie. La filosofia degli ibridi deve
essere quindi abbracciata con tutte le cautele del
caso. Il Girfalco bianco è il sogno di molti
falconieri, ma solo in pochi possono acquistarne uno
(possibilmente una femmina che è notevolmente più
grossa del maschio); la tecnica dell’ibridazione
permette di produrre con poca spesa (cioè avendo
solo un maschio di Girfalco) degli ibridi GirxSacro,
tanto per fare un esempio, che avranno un colore
bianco quasi come il Girfalco puro, un peso
sicuramente superiore a quello del Sacro (ma
inferiore a quello del Girfalco puro appunto perché
le caratteristiche dell’ibrido sono una via di mezzo
tra i due genitori) ma allo stesso tempo un prezzo
ben più abbordabile rispetto ad un Girfalco puro; e
questo senza considerare le eventuali implicazioni
etologiche, perché il GirxSacro è un ottimo ibrido
da caccia che offre delle performance migliori del
Sacro puro in alcune particolari circostanze.
Dall’esempio appena riportato si capisce facilmente
il perché della produzione di ibridi di falchi.
C'è un
ultimo punto da affrontare e cioè il fatto che un
ibrido potrebbe essere ritenuto in qualche modo
innaturale o non sportivo. Certamente non si può
dire che un ibrido sia poco naturale quando poi
vengono fatti volare per la falconeria delle specie
pure come i Pellegrini per es. usando la telemetria
o trasportando i rapaci in macchina con il cappuccio
o tenendoli in città o su selvaggina di rilascio o
su un pallone aerostatico, anche questi modi di fare
falconeria allora sono tutti “innaturali”. Per
quanto riguarda la non sportività bisogna dire che
esiste sempre un equilibrio tra predatore e preda e
tale bilancio viene mantenuto dal falconiere anche
usando un ibrido. Per chiarire meglio il concetto di
sportività ricorriamo ad un esempio: in Inghilterra
ci sono delle aree molto ricche di prede e molto
adatte alla caccia con il Pellegrino ma in esse è
vietata qualsiasi forma di caccia così i falconieri
sono costretti ad usare i falchi pellegrini in zone
non adatte a tale rapace a causa dell’ eccessiva
presenza di vegetazione. In tali aree dunque un
Falco pellegrino non potrà esprimersi al meglio, non
potrà cioè cacciare in picchiata e non potrà
inseguire la preda fino all'ultimo se essa si
nasconde tra la vegetazione. In una tale circostanza
dunque sarà effettivamente più redditizio l'uso di
un ibrido per esempio tra Falco sacro e Falco
pellegrino, esso infatti possedendo i geni di
entrambe queste due specie potrà sia cacciare in
picchiata sia catturare le prede anche tra la
vegetazione. L’ibridologia permette alla falconeria
moderna di adattarsi alla società moderna.GLI IBRIDI SELVATICI
Anche
in natura gli ibridi non sono rarissimi, sia nei
Mammiferi quanto negli Uccelli (quelli di Fringuello
x Peppola sono noti da tempo ma si sono osservati
anche ibridi tra Rondine x Balestruccio e i
"cacciatori di valle" sanno bene che è abbastanza
frequente la cattura di anatre che risultano nate
allo stato libero da individui di Specie differenti
per es. Moriglione x Germano, Morette, Fischione,
ecc.). In generale l’ibridazione naturale avviene
per le specie che non hanno evoluto dei
comportamenti corteggiativi complessi come per es. i
Tetraonidi, e i Trochilidae, oppure è frequente in
popolazioni molto numerose (infatti Mayr e Short,
1970 non riuscirono a trovare ibridi in nessuna
specie rara nordamericana). Anche per quanto
riguarda i rapaci si conoscono degli ibridi
naturali. Bisogna considerare che tali ibridi di
solito possono nascere perché le specie coinvolte
sono fenotipicamente molto simili e se a questo si
aggiunge che vi possono essere delle interferenze
nel normale imprinting sessuale oltre che delle
eccezioni e delle anomalie (la biologia non è
matematica) ecco che si possono spiegare alcune di
queste ibridazioni, a maggior ragione se poi le
specie coinvolte sono simili anche dal punto di
vista filogenetico per cui non si presenteranno
delle grosse “barriere riproduttive”. Si conoscono
ibridi tra Nibbio bruno (Milvus migrans) e
Poiana comune (Buteo buteo) (Corso & Forsman,
1997), tra Avvoltoio nero e Grifone (McIlhenny,
1937), tra Pellegrino e Falco della prateria (Falco
mexicanus) (Oliphant, 1991) ecc. (si veda la tabella
sotto per un elenco, anche se parziale, degli ibridi
naturali tra rapaci). Oliphant (1991) ha descritto
una ibridazione naturale tra Falco pellegrino e
Falco della prateria, che mi sembra utile citare
perché è possibile, in parte, spiegarla. Infatti si
è visto che il maschio di Pellegrino che è riuscito
ad accoppiarsi con la femmina di Falco della
prateria (e ciò grazie al fatto che le due specie
hanno comportamenti corteggiativi molto simili)
aveva un anellino del Peregrine Fund che indicava
che era uno degli individui rilasciati nei programmi
di reintroduzione portati avanti in quegli anni.
Probabilmente, allora, questa ibridazione naturale
può essere spiegata col fatto che molti Pellegrini
nati in cattività e poi rilasciati dal Peregrine
Fund sono stati fatti allevare da genitori adottivi
(cross-fostering) Falchi della prateria (allo scopo
di lasciare le femmine di Pellegrino libere di
deporre altre uova); questi Pellegrini allora
avevano acquisito un imprinting sessuale sulla
specie Falco della prateria, e ciò giustifica la
loro ibridazione naturale. Mi sembra giusto però
ricordare che ciò è avvenuto in anni in cui non si
sapeva ancora niente degli effetti dell’imprinting
sulla scelta sessuale.’ETICA DEGLI IBRIDI
Per
quanto riguarda la produzione di ibridi artificiali
da usare nella falconeria potrebbe sorgere una
questione: visto che a volte capita che il
falconiere (per un suo errore o per circostanze
dovute al caso) può perdere il controllo del suo
Falco, quali probabilità ci possono essere che se
questo Falco è un ibrido, esso possa reincrociarsi
con un Falco selvatico? E se ciò avvenisse, che
rischi ci sono che gli ibridi ottenuti da questa
riproduzione possano alterare il pool genico della
popolazione selvatica? A questa domanda non c'è una
risposta completa. Ci sono due aspetti da tenere in
considerazione: un ibrido ed un rapace selvatico
possono riprodursi (in altre parole essi sono
sufficientemente geneticamente compatibili)? E, si
può formare una coppia tra loro (cioè possono
riconoscersi l’un l’altro come potenziali partner)?
Per esempio il solo grosso falcone che abbiamo in
Italia è il Pellegrino. Un ibrido tra "Shaheen"e
Pellegrino accoppiandosi con un Pellegrino, è
biologicamente capace di dar vita ad una progenie
probabilmente anch’essa fertile (ricordiamo che si
tratta di incroci tra sottospecie, che quindi hanno
un alto grado di fertilità), ma un ibrido tra
Pellegrino e Falco del deserto (per es. GyrxPell)
difficilmente riuscirà a dare progenie se si
accoppia con un Pellegrino, perché il Pellegrino
appartiene ad un sottogenere (Rhincodon) diverso dal
sottogenere cui appartengono i Falchi del deserto
(Hierofalco). In Inghilterra non ci sono Lanari ma
quelli che vengono usati per la falconeria sono
stati a volte persi però non si è verificato mai un
caso di incrocio tra Lanari e Pellegrini. Si sa
comunque che l' imprinting sui genitori influenza in
maniera critica la futura selezione del compagno.
Così per esempio un Falco sacro allevato da un
Pellegrino più facilmente accoppiarsi con un
Pellegrino selvatico, ma in tal caso sarà
probabilmente il Pellegrino selvatico a rifiutare
l’accoppiamento, riconoscendo il Sacro come una
specie estranea (non conspecifica) perché ha un
imprinting sessuale naturale essendo stato allevato
dai suoi genitori Pellegrini.
Molti
Paesi hanno preso coscienza del pericolo (sebbene
remoto) che gli ibridi potrebbero arrecare alle
popolazioni selvatiche. Molte associazioni di
falconeria si sono date delle regole finalizzate a
ridurre questi rischi, per esempio anche bandendo
completamente l’uso degli ibridi tra i loro membri.
In Medioriente dove gli ibridi di GyrxSacro sono
usati molto comunemente nella falconeria, si sta
cercando di evitare qualsiasi possibilità di
inquinamento genetico della popolazione selvatica di
Falco sacro ricorrendo alla sterilizzazione di tali
ibridi. Esistono varie metodologie che praticamente
annullerebbero qualsiasi rischio di inquinamento dei
pool genici delle popolazioni selvatiche:
l’imprinting sull’uomo o su specie che non si
trovano allo stato selvatico in quella regione,
l’eventuale sterilizzazione degli ibridi (a parte
che gli ibridi spesso sono già poco fertili o
completamente sterili per loro fisiologia),
l’obbligo dell’uso delle apparecchiature da
radiotracking, sono solo alcune, se poi a queste
aggiungiamo che è molto difficile che un ibrido
sfuggito alla cattività possa riuscire ad
accoppiarsi, soprattutto se non è ancora addestrato
per la falconeria, in tal caso morirebbe prima di
potere riprodursi, e che gli eventuali ibridi nati
da questi incroci con gli individui selvatici
saranno probabilmente sterili, ogni pericolo sarà
eliminato. La tecnica per gestire correttamente i
rapaci ibridi è di imprintarli sull'uomo o su una
specie che non si trova allo stato selvatico nella
regione in cui viene allevata questa specie o viene
usata per la falconeria. Il solo esempio, in
Inghilterra, in cui c'è un serio rischio di incrocio
è l'accoppiamento tra la Poiana codarossa e la
Poiana comune. Anche su questo fenomeno ci sono
pochissimi dati e non si conoscono finora dei casi
che dimostrino che ciò sia avvenuto. Ma
nell'eventualità che ciò accada su scala sufficiente
da creare preoccupazioni è meglio sterilizzare o
effettuare un "malimprinting" delle codarossa, ma
non c'è bisogno di fermare l'uso di specie esotiche
nella falconeria come per esempio le Poiane di
Harris e le codarossa stesse. Alcuni recenti studi
hanno mostrato che le varie specie di falconi hanno
in comune una gran parte dei loro geni così come
l'uomo ha in comune circa il 95% del suo materiale
genetico con altri primati. Il nostro intero
concetto di specie sta cambiando e l'edificio
tassonomico sta crollando. Così sarebbe una follia
tentare di giungere a delle conclusioni con i dati
di cui siamo in possesso fino ad oggi.
La
base del problema dunque, da quanto visto, non è
tanto la presenza di ibridi ma più che altro
l'inquinamento del pool genico delle specie
selvatiche. Questo è un problema mondiale e già
esistono numerosi casi di inquinamento genetico per
esempio gli incroci tra gatti domestici e gatti
selvatici o tra cani inselvatichiti e lupi. Quindi è
giusto che i geni selvatici rimangano tali e non
vengano inquinati ma questo implica anche che l'
habitat delle popolazioni rimanga naturale e non si
inquini! Ciò però non è facile.
L'inquinamento genetico non è comunque una fattore
così grave. Infatti a volte l'introduzione di geni
esterni attraverso l’out-breeding può aiutare le
popolazioni selvatiche ad adattarsi meglio a nuove
(e purtroppo attuali) variazioni ambientali e
climatiche. In nord America, per esempio, il
Peregrine Fund ha rilasciato al posto del Falco
pellegrino della ssp. anatum, che si era
quasi estinta, varie ssp. di Pellegrini nella
speranza che si selezionasse naturalmente e
rimanesse solo quella più adatta alle circostanze
odierne. E questo potrebbe essere un es. di
inquinamento dei geni selvatici, solo all’apparenza
però; si è, infatti, scelto di operare appositamente
così perché il pool genico della popolazione
selvatica da restaurare con le reintroduzioni si era
drammaticamente ridotto a causa della riduzione
notevole del numero di individui (e del conseguente
inbreeding, della deriva genetica ed eventuali
effetti a “collo di bottiglia”) che la compongono.
Introdurre geni estranei è servito ad allargare la
variabilità genetica della popolazione, e come si sa
la variabilità è la base dell’evoluzione e delle
capacità adattative di una popolazione.
L’inquinamento del pool genico di una popolazione
può avvenire anche in cattività, e questa, di
solito, è una cosa da condannare aspramente. Spesso
gli allevatori privati sfruttano per la riproduzione
tutti gli individui di cui possono disporre, visto
che i rapaci in cattività sono costosi, allora se
l’allevatore dispone di una femmina di Falco
peregrinus brookei e di un maschio Falco
peregrinus peregrinus, molto spesso non esiterà
ad accoppiarli, pur trattandosi di esemplari di ssp.
diversa, e produrrà quindi degli ibridi
intraspecifici. Questi ibridi non servono a niente,
di solito (una eccezione è per esempio quella del
Peregrine Fund illustrata sopra). Anzi, danneggiano
il pool genico delle popolazioni in cattività.
Questi incroci ottenuti in F1 (nel caso dell’es.
sopra F.p.bookei x F.p.peregrinus)
verranno reincrociati ulteriormente nelle successive
generazioni e si perderà la traccia di tutti gli
incroci e reincroci fatti. Alla fine si otterranno
sì Falchi pellegrini ma che non si sa di che ssp.
siano e raramente potranno avere un utilizzo al di
fuori della riproduzione in cattività e della
falconeria. E’ dunque da condannare questo modus
operandi, bisognerebbe rendere obbligatori i
pedigree ed i test genetici per tutti i rapaci
prodotti in cattività (questo aiuterebbe anche ad
evitare problemi di inbreeding e a gestire meglio la
legalità degli esemplari).
Adesso
viene spontanea una domanda: un ibrido se viene
perso può sopravvivere allo stato selvatico?
Certamente un ibrido ucciderà molte prede quando
viene usato per la falconeria con risultati
addirittura migliori di quelli ottenuti dalla specie
pura. Allora si può pensare che un tal ibrido
potrebbe continuare a vivere allo stato selvatico. E
così esso può sopravvivere come un singolo individuo
per tutta la sua vita ma non avrà la possibilità di
diffondere il suo genoma perché gli risulterà
difficile accoppiarsi. I rapaci da falconeria in
generale (ibridi e non) non ancora addestrati
avranno invece una bassissima possibilità di poter
sopravvivere allo stato selvatico, e allora è
essenziale che il falconiere prenda tutte le
possibili precauzioni per prevenire la perdita di un
rapace soprattutto se non adeguatamente addestrato
(obbligo di uso del radiotracking ecc.).
Ultimo
argomento da affrontare, visto che stiamo parlando
di morale è se sia moralmente giusto produrli: a
questo proposito ci si deve chiedere se sia giusta,
allora, la produzione di tante razze nuove di cani o
gatti domestici. A parte questo bisogna dire che un
ibrido non si rende conto di essere tale, non ha
coscienza, la coscienza è una prerogativa esclusiva
dell’Homo sapiens. L’importante è che venga
tenuto nel massimo benessere fisico e psichico ed
evitandogli qualsiasi sofferenza, ma a questo punto
il problema si estende a tutti gli animali in
cattività…
Allo
scopo di illustrare in pratica le particolari
caratteristiche degli ibridi, ho raccolto alcuni
dati bibliografici su alcuni tipi di ibridi oggi
molto usati in falconeria.
A)
IBRIDO PELLEGRINO X PRATERIA
PESO
DI VOLO: Maschio 500-600 g Femmina 750-870 g
Il
primo di questi ibridi è nato negli USA nei primi
anni ’80. Questo ibrido è più pesante di un Falco
della prateria ma più leggero, nel peso di volo, di
una femmina di Pellegrino. E’ un ibrido molto
aggressivo, vivace e coraggioso. Le femmine sono
molto adatte alla caccia al fagiano, alla pernice,
alla starna ed alle anatre e si adattano bene alle
prede di basso volo. Con il loro più leggero carico
alare e con la loro coda più lunga rispetto al
Pellegrino questi ibridi sono molto agili e possono
picchiare in modo molto leggero e rapido. L’
influenza genetica dei due tipi di genitori permette
a questo ibrido di poter catturare le prede
attaccandole da alte posizioni aeree, in picchiata,
tecnica in cui questo tipo di ibrido eccelle. Sono
tipiche le picchiate rapide e potenti, fatte con un
coraggio che manca nel Falco della prateria. Questo
Falco è molto manovrabile e può avere delle
performance straordinarie uccidendo prede di basso
volo (come le pernici) dopo una picchiata da una
posizione altissima e con la tipica astuzia del
Falco della prateria che trattiene la preda prima
che essa tocchi il suolo. Dal punto di vista del
temperamento questo ibrido è più fiero del
Pellegrino ma lo è meno di un Falco della prateria.
E’ molto adattabile alle condizioni atmosferiche
ventose, ma a causa del loro basso carico alare il
vento eccessivamente forte può impedirgli di
effettuare delle picchiate veloci. In condizioni di
forte vento quindi un ibrido di Pellegrino x
Girfalco o un semplice Pellegrino otterranno dei
risultati migliori. I maschio dell’ ibrido Prateria
x Pellegrino hanno anch’ essi molto talento e sono
adatti a tutte le specie di uccelli terricoli
comprese le starne. Possono catturare anche varie
specie di Corvidi, grazie al fatto che questo ibrido
è molto paziente e persistente nella caccia. I
maschi hanno molta agilità, un carattere derivante
dall’ influenza del Falco della prateria e sono
capaci di lanciarsi dentro la fitta vegetazione allo
scopo di catturare le loro prede.
B)
IBRIDO GIRFALCO X PELLEGRINO
PESO
DI VOLO:maschio 650-750 g femmina 1100-1170 g
Il
peso di volo sopraindicato è tipico dell’ ibrido del
Girfalco con il F. peregrinus peregrinus
e non con altre sottospecie più piccole di
Pellegrino. L’ibrido Girfalco x Pellegrino è di
grosse dimensioni e molto potente. Le femmine sono
capaci di catturare tutti i tipi di uccelli
terricoli e non solo. Questo tipo di ibrido ha
mostrato di avere molti tratti in comune con il
Girfalco soprattutto nell’ atteggiamento. Come i
Girfalchi esso ha una capacità di accelerazione
molto potente e caccia spesso con la tecnica dell’
inseguimento (“cul levè”) e del volo di attesa
(“waiting on flight”). Infatti il vantaggio di
questo ibrido rispetto al Girfalco puro è che esso è
più ubbidiente e coopera di più nel volo di attesa
col falconiere. Come i Girfalchi esso richiede un
grande sforzo per rimanere allenato (soprattutto le
femmine) a causa del loro grosso peso corporeo
comparato con quello dei Pellegrini puri. Comunque
le femmine di GyrxPell sono molto adatte a volare in
presenza di venti molto forti grazie alla loro
potenza di volo, a differenza dei Pellegrini che
invece volano male. Questo ibrido è molto simile al
Girfalco anche per quanto riguarda la capacità di
manovrare in volo, esso infatti possiede una potente
capacità di accelerazione ed una notevole velocità
durante la caccia grazie alla sua notevole massa
muscolare. Questo tipo di ibrido ha bisogno di
lavorare in campi moto aperti. Grazie alla capacità
di manovra tipica del Girfalco esso può esplorare
larghe aree di suolo così che fagiani, anatre e
starne sono per lui delle facili prede. Questo tipo
di ibrido infine può essere utilizzato anche per la
caccia ai Corvidi.

Ibrido Girfalco nero x Pellegrino
C)
IBRIDO GIRFALCO X SACRO
PESO
DI VOLO: Maschio 850-950 g Femmina1200-1250 g
Di
tutti gli ibridi con il Girfalco questo è il più
simile al Girfalco. Il Girfalco ed il Falco sacro
sono molto simili sia fisicamente sia mentalmente, e
la loro combinazione produce un tipo di Girfalco più
leggero e più vivace. Le femmine di questo tipo di
ibrido sono capaci di catturare grosse prede e la
loro velocità è notevole. Esse possono anche
catturare le lepri grazie alla loro potenza. Come il
Girfalco, questo tipo di ibrido ha bisogno di
giornate ventose per poter cacciare al meglio,
inoltre esso possiede una grande apertura alare
(comparata a quella del Falco sacro) la quale gli
permette di essere usato anche in giornate
tempestose. Sia il maschio sia la femmina di questo
tipo di ibrido possono essere utilizzate con
successo nella caccia alle prede terricole. La
maggiore area alare ed il conseguente minor carico
alare consentono al GyrxSacro di raggiungere altezze
elevatissime. All’origine, l’ibrido GyrxSacro è
stato prodotto per consentire ai falconieri arabi di
utilizzare falconi di enorme dimensione in ambiente
desertico, a cui il Girfalco puro non è adattato. La
selezione odierna dei GyrxSacro punta verso
l’ottenimento di individui di mole sempre maggiore e
colore sempre più puro (bianco o nero) mantenendo un
buon adattamento a temperature elevate e clima secco
(situazione tipicamente desertica).

Ibrido Girfalco bianco x Sacro
La
tabella seguente illustra alcuni degli ibridi
naturali ed artificiali di cui sono venuto a
conoscenza tramite la bibliografia. Se ne conoscete
altri, per favore segnalatemeli. Come si vede negli
Strigiformes i casi di ibridazione naturale ed
artificiale sono rarissimi. In particolare non
esistono ibridi artificiali proprio perché non
avrebbero alcuna utilità; anche se sicuramente si
può affermare l’esistenza in cattività di ibridi (o
meglio incroci) tra varie sottospecie, per es. sono
a conoscenza di ibridi tra Bubo bubo bubo e
Bubo bubo sybiricus (Gufo reale europeo e
Gufo reale siberiano).
Dalla
tabella ho omesso tutti i tri- e quadri- ibridi così
come tutti gli F2 e gli R2 di rapaci in cattività di
cui sono a conoscenza, la lista risulterebbe troppo
lunga
|
|
IBRIDI NATURALI |
IBRIDI ARTIFICIALI
|
|
FALCONIFORMES |
Chatartes aura x coragyps auratus
Aegypius monachus x Gyps fulvus
Gyps ruppellii x Gyps africanus
Milvus milvus x Milvus migrans
Accipiter genitlis x Buteo buteo
Buteo buteo x Buteo jamaicensis
Milvus migrans x Buteo buteo
Falco peregrinus x Falco mexicanus
Falco cherrug x Falco peregrinus
Falco cherrug x Falco biarmicus
Falco rusticolus x Falco cherrug
Falco tinnunculus x Falco naumanni
Accipiter fasciatus x Accipiter
novaehollandiae
|
Falco peregrinus x Falco biarmicus
Falco peregrinus x Falco cherrug
Falco peregrinus x Falco columbarius
Falco peregrinus x Falco rusticolus
Falco peregrinus x Falco mexicanus
Falco peregrinus x Falco sparverius
Falco peregrinus x Falco deiroleucus
Falco peregrinus x Falco rufigularis
Falco cherrug x Falco rusticolus
Falco cherrug x Falco biarmicus
Falco rusticolus x Falco sparverius
Falco rusticolus x Falco columbarius
Falco rusticolus x Falco mexicanus
Parabuteo unicinctus x Buteo jamaicensis
Accipiter cooperii x Parabuteo
unicinctus
Aquila chrisaetos x Aquila rapax |
|
STRIGIFORMES |
Otus asio x Otus kennicotti |
|
BIBLIOGRAFIA
BIBLIOGRAFIA
1.
Baptiste, S. and D. Jamieson. 1978. [Peregrine
falcon--prairie falcon hybridization experiments.].
Unpubl.
Rep.
Reno, Nevada. 5 Pp..
2.
Boyd, L. and N. Boyd. 1975. Hybrid falcon: The
sharie at fledging. Hawk Chalk 14:53-54.
3.
Cade, T.J. 1986. Propagating diurnal raptors in
captivity: a review. International Zoo Yearbook
24/25:1-20.
4.
Cade, T.J. and J.D. Weaver. 1976. Gyrfalcon X
peregrine hybrids produced by artificial
insemination. NAFA 15:42-47.
5.
Corso, A. and D. Forsman. 1997. Hybrids between
black kite and common buzzard. Alula
3:44-45.
6.
Corso, A. and R. Gildi. 1998. Hybrids of black
kite and common buzzard in Italy in 1996. Dutch
Birding 20:226-233.
7.
Dark, S.J., R.J. Gutiérrez and G.I. Gould Jr.
1998.
The barred owl (Strix varia) invasion in
California. Auk 115:50-56.
-
Ellis, D. H. 1995. What is Falco altaicus
Menzebier?
J. Rap. Res. 29 :15-25
9.
Fairclough, K. 1919. The pallid harrier in Orkney.
Birding World :253-255.
10.
Fleischer, R.C. 1998. Genetics and avian
conservation. pp. 29-47. In J.M. Marzluff and
R. Sallabanks [eds.] . Avian conservation: research
and management. Island Press, Washington, D.C. (Some
papers presented at a symposium held at the 1996
joint meeting of the AOU and RRF)
11.
Forsman, E. 1995. Spotted owl monitoring protocols
for demographic studies. USDA, Forest Service,
Pacific Norwest Research Station, Portland, Oregon.
-
Grant, P. R. and B. R. Grant. 1992.
Hybridization of bird species. Science
256:193-197
13.
Haig, S.M. and J.C. Avise. 1995. Avian
conservation genetics. pp. 160-189. In J.C.
Avise and J.L. Hamrick [eds.] . Conservation
genetics: case histories from nature. Chapman &
Hall, New York.
14.
Hamer, T.E., E.D. Forsman, A.D. Fuchs and M.L.
Walters. 1994. Hybridization between barred and
spotted owls. Auk 111:487-492.
15.
Hamer, T.E., E.D. Forsman, A.D. Fuchs and
M.L. Waters. 1993. Abstract: Hybridization between
barred and spotted owls. Journal of Raptor Research
27:72. (Abstr. presentation, annu. meet. Raptor
Research Foundation, Inc., Bellevue, Washington,
11-15 November, 1992)
16.
Hardaswick, V.J. and D.G. Smith. 1981. Peregrine X
prairie hybrids: production & characteristics.
Journal of the North American Falconers'
Association 20:4-11.
17.
Hardaswick, V.J., D.G. Smith and T.J. Cade. 1984.
Growth and behavioral development of peregrine x
prairie falcon hybrids. NAFA 23:13-21.
18.
Jefferson, B. 1994. Successful hybridization of
common raven and American crow. Ontario Birds
12:32-35.
19.
Keyser, C., B. Ludes, H. Pfitzinger and P. Mangin.
1995. Detection of sex-specific DNA fingerprint
bands in the peregrine falcon (Falco peregrinus)
using non radioactive 33.6 and 33.15 human probes.
Ethology Ecology & Evolution 7:355-361.
20.
Lány, P., I. Rychlík, J. Bárta, J. Kundera and I.
Pavlík. 1999.
Salmonellae in one falcon breeding facility in the
Czech Republic during the period 1989-1993. Veterinarni
Medicina - Czech 44:345-352.
21.
Levy, S. 1999. Owl vs. owl. Natural History
108:28-32.
22.
Lindsay, D.S., D.S. Zarlenga, H.R. Gamble, F.
Al-Yaman, P.C. Smith and B.L. Blagburn. 1995.
Isolation and characterization of Trichinella
pseudospiralis garkavi, 1972 from a black
vulture (Coragyps atratus). Journal of
Parasitology 81:920-923.
23.
Linthicum, J. 1988. Peregrine falcon
monitoring, nest management, hack site, and
cross-fostering efforts, 1988. Unpubl. Rep. Santa
Cruz Predatory Bird Research Group, University of
California, Santa Cruz. 75 Pp..
-
Mayr, E. 1970. Populations, species and
evolution. Harvard Univ. Press, Cambridge, MS
USA.
25.
McIlhenny, E.A. 1937. A hybrid between turkey
vulture and black vulture. Auk
54:384.
26.
Mindell, D.P. 1983. Harlans hawk (Buteo
jamaicensis harlani): a valid subspecies. Auk
100:161-169.
27.
Morris, J. and R. Stevens. 1970. Successful
cross-breeding of a peregrine tiercel and a saker
falcon. Captive Breeding of Diurnal Birds of Prey,
R.E. Kenward and G.F. Jolly, eds., Bol 1., No. 1.,
Brit. Falconers Club and the Hawk Trust. pp 55-7.
28.
Murphy,
C.J., T.J. Kern, E. Loew, N.C. Buyukmihci, R.W.
Bellhorn, A. De Lahunta, W. Heck and D.L. Graham. 1985.
Retinal dysplasia in a hybrid falcon. Journal of
the American Veterinary Medical Association
187:1208-1209.
29.
Nagy, T. 1998. Data on distribution of white-brested
barn owls (Tyto alba). Aquila
103-104:149-150.
30.
Norman, J., P. Olsen and L. Christidis. 1998.
Molecular genetics confirms taxonomic affinities of
the endangered Norfolk Island boobook owl Ninox
novaeseelandiae undulata. Biological
Conservation 86:33-36.
31.
Oliphant, L.W. 1991. Hybridization between a
Peregrine Falcon and a Prairie Falcon in the wild.
Journal of Raptor Research 25:36-39.
32.
Olsen, P. 1990. Social behavior. pp. 108-123.
In I. Newton [ed.] . Birds of prey. Facts on
File, New York.
33.
Palmer, R.S. 1988. Family Cathartidae new
world condors and vultures; family Accipitridae
(first part) osprey, kites, bald eagles and allies,
accipiters, harrier, Buteo allies. pp. 355-378.
In R.S. Palmer [ed.] , Vol. 4. Handbook of North
American Birds. Yale University Press, New Haven,
CT.
34.
Parrish, J.R. and C.M. White. 1997.
Abstract: Comments on hybridization in raptors.
Journal of Raptor Research 31:296-298. (Abstr.
presentation, annu. meet. {IRaptor Research
Foundation, Inc.}, Gainsville, Florida, 1986)
35.
Ruos, J.L. 1978. Raptor hybridization--a
review of the occurence, significance, and
regulation of activities permitted under special
purpose permit. Unpubl. rep. U.S. Fish and Wildlife
Service, Washington D.C. 12pp.
36.
Rychlík, I., O. Kubícek, V. Holcák, J. Bárta and I.
Pavlík. 1994.
DNA fingerprinting in Falconidae. Veterinarni
Medicina - Czech 39:111-116.
37.
Schlorff, R.W. 1981. Captive raptor breeding
program. Job progress report, nongame wildlife
investigations, California dept. of Fish & Game,
Sacramento, California. 12pp.
38.
Schmutz, S.M. and L.W. Oliphant. 1987. Chromosome
study of peregrine, prairie, and gyrfalcons with
implications for hybrids. Journal of Heredity
78:388-390.
-
Sibley, C. G., 1957. The evolutionary and
taxonomic significance of sexual selection and
hybridization in birds. Condor 59:166-191
40.
Stevens, R. 1972. B.P.I.E. No. 27.
Peregrine Falcon-Saker cross. Raptor Research
6:18-21.
41.
Stevens, R. 1972. B.P.I.E. No. 49.
Peregrine Falcon x Saker hybrid. 1
p..
42.
Stevens, R. 1974. B.P.I.E. No. 127.
Peregrine Falcon x Saker hybrid. 4
pp..
43.
Stevens, R. 1974. B.P.I.E. No. 96.
Peregrine Falcon x Saker hybrid. 3
pp..
44.
Walton, B.J. 1981. Captive breeding 1981. Unpubl.
rep. submitted to California Dept. of Fish and Game,
Non-game Wildlife Management Section, Sacramento,
California.
41 pp.
45.
Walton, B.J. 1987. [California Raptor Conservation
Committee recommended policy on hybridization of
captive raptors.]. Letter to P.B. Bontadelli Dated
June 17, 1987. 2 Pp..
46.
Walton, B.J. 1987. [California Raptor Conservation
Committee recommended policy on hybridization of
captive raptors.]. Letter to P.B. Bontadelli Dated
June 17, 1987. 2 Pp..
47.
Woinarski, J.C.Z. 1998. Research and
management for the conservation of birds in
Australia. pp. 383-393. In J.M. Marzluff and
R. Sallabanks [eds.] . Avian conservation: research
and management. Island Press, Washington, D.C. (Some
papers presented at a symposium held at the 1996
joint meeting of the AOU and RRF)
48.
Wolhuter, B.R. and F. Kish. 1970. Courtship
display observed between two species of buteos.
Wilson Bulletin 82:96-97.
Volete
Approfondire questo argomento?
Se volete
imparare di più le tecniche e approfondire i
concetti di questa pagina web,
consultate i
nostri prodotti multimediali, libri e CD
|
|
